Concept 7.4: Meiosis Halves the Nuclear Chromosome Content and Generates Diversity
In Concept 7.l we described the role and importance of meiosis in sexual reproduction. Now we will see how the orderly and precise generation of haploid cells is accomplished.
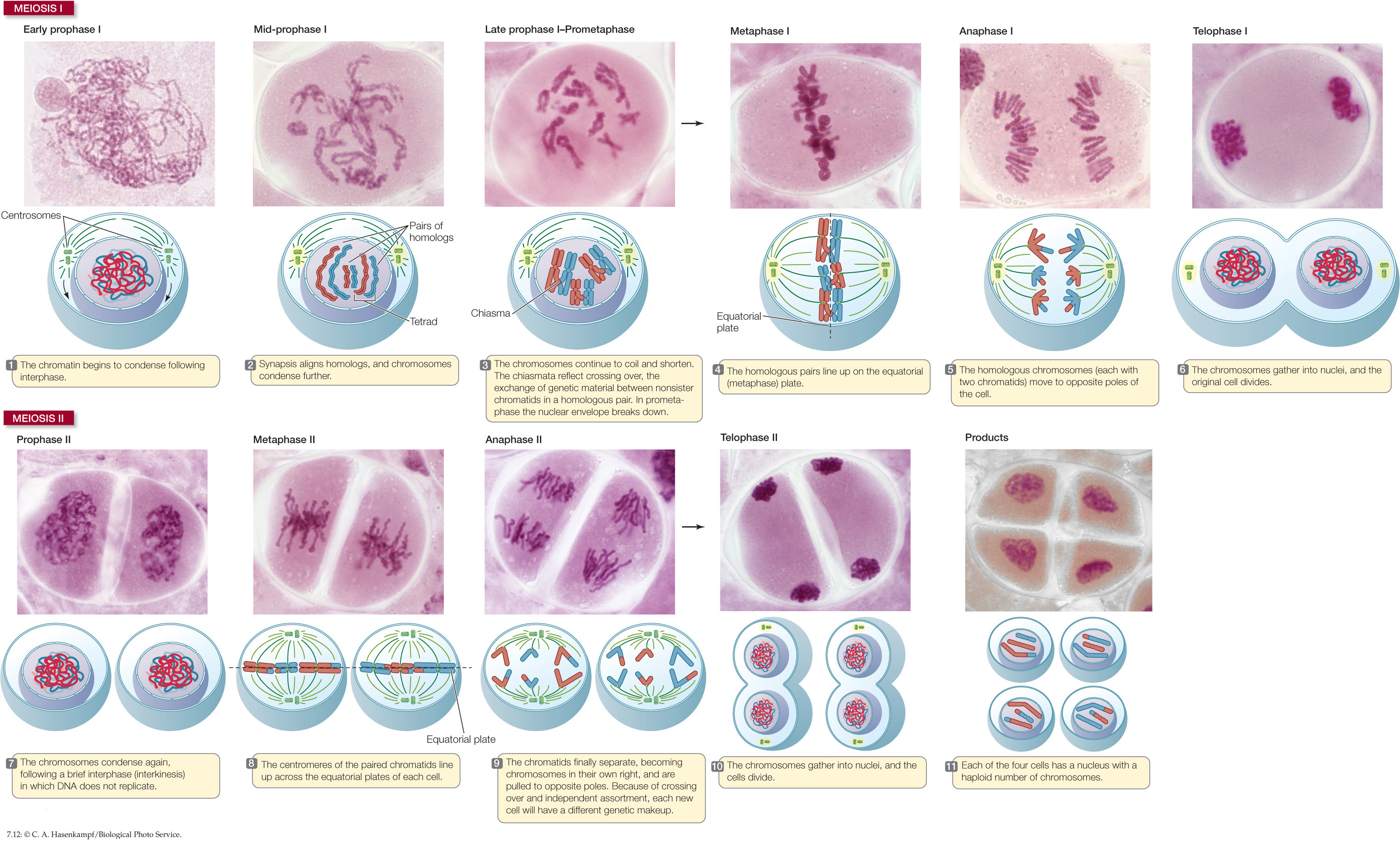
Meiosis consists of two nuclear divisions that reduce the number of chromosomes to the haploid number. Although the nucleus divides twice during meiosis, the DNA is replicated only once. Unlike the products of mitosis, the haploid cells produced by meiosis are genetically different from one another and from the parent cell. FIGURE 7.11 compares the two processes.

To understand the process of meiosis and its specific details, it is useful to keep in mind the overall functions that meiosis has evolved to serve:
- To reduce the chromosome number from diploid to haploid
- To ensure that each of the haploid products has a complete set of chromosomes
- To generate genetic diversity among the products (gametes)
The events of meiosis are illustrated in FIGURE 7.12. In the rest of this section we will discuss some of the key features that distinguish meiosis from mitosis.
Go to ACTIVITY 7.4 Images of Meiosis
PoL2e.com/ac7.4

Go to ANIMATED TUTORIAL 7.2 Meiosis
PoL2e.com/at7.2
Meiotic division reduces the chromosome number
As noted above, meiosis consists of two nuclear divisions, meiosis I and meiosis II. Two unique features characterize meiosis I:
- Homologous chromosomes come together and line up along their entire lengths. No such pairing occurs in mitosis.
- The homologous chromosome pairs separate, but the individual chromosomes, each consisting of two sister chromatids, remain intact. (The chromatids will separate during meiosis II.)
Like mitosis, meiosis I is preceded by an interphase with an S phase, during which each chromosome is replicated. As a result, each chromosome consists of two sister chromatids. At the end of meiosis I two nuclei form, each with half of the original chromosomes (one member of each homologous pair). Since the centromeres did not separate, these chromosomes are still double—composed of two sister chromatids. The sister chromatids are separated during meiosis II, which is not preceded by DNA replication. As a result, the products of meiosis I and II are four cells, each containing the haploid number of chromosomes. But these four cells are not genetically identical.
Crossing over and independent assortment generate diversity
A diploid organism has two sets of chromosomes (2n): one set derived from its male parent, the other from its female parent. As the organism grows and develops, its cells undergo mitotic divisions. In mitosis, each chromosome behaves independently of its homolog, and its two chromatids are sent to opposite poles during anaphase. Each daughter nucleus ends up with an identical set of 2n chromosomes. In meiosis, things are very different (see Figure 7.11).
141
142
An important consequence of meiosis is that the four resulting cells differ from one another genetically. The shuffling of genetic material occurs by two processes: crossing over and independent assortment.
143
Crossing Over
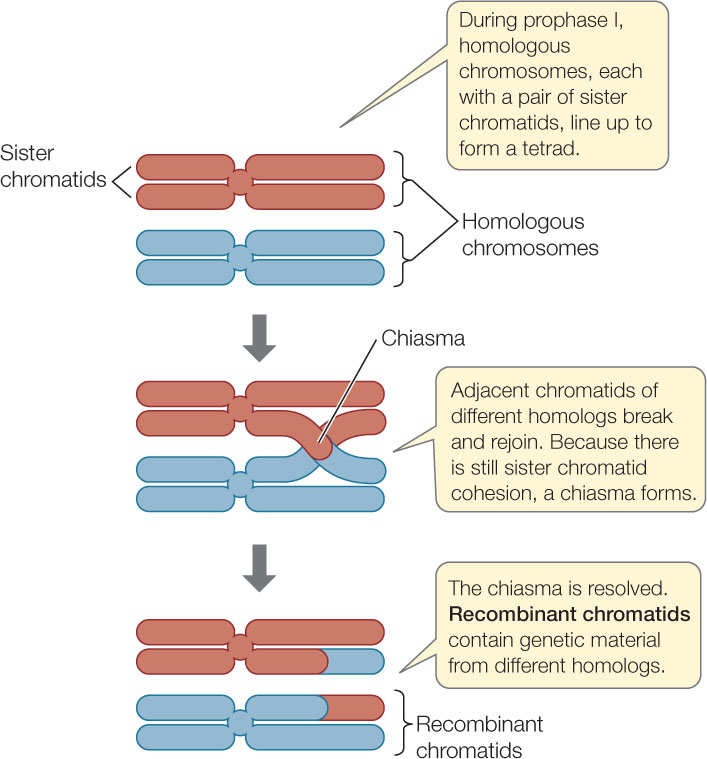
Meiosis I begins with a long prophase I (the first three panels of Figure 7.12), during which the chromosomes change markedly. The homologous chromosomes pair by adhering along their lengths in a process called synapsis. (This does not happen in mitosis.) This pairing process lasts from prophase I to the end of metaphase I. The four chromatids of each pair of homologous chromosomes form a tetrad, or bivalent. For example, in a human cell at the end of prophase I there are 23 tetrads, each consisting of four chromatids. The four chromatids come from the two partners in each homologous pair of chromosomes.
Throughout prophase I and metaphase I, the chromatin continues to coil and compact and the chromosomes become more condensed. At a certain point, the homologous chromosome pairs appear to repel each other, especially near the centromeres, but they remain attached. The X-shaped attachment points are called chiasmata (singular chiasma, “cross”):
144
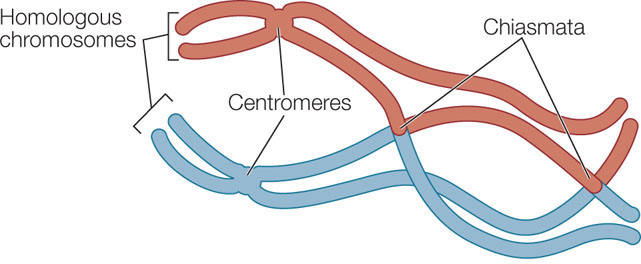
A chiasma is a point where genetic material is exchanged between nonsister chromatids on homologous chromosomes—a process called crossing over (FIGURE 7.13). Any of the four chromatids in the tetrad can participate in this exchange, and a single chromatid can exchange material at more than one point along its length. Crossing over occurs shortly after synapsis begins, but chiasmata do not become visible until later, when the homologs are repelling each other. Crossing over results in recombinant chromatids, and it increases genetic variation among the products of meiosis by reshuffling genetic information between homologous chromosome pairs. In Concept 8.3 we will explore further the genetic consequences of crossing over.
Mitosis seldom takes more than an hour or two, but meiosis can take much longer. In human males, the cells in the testis that undergo meiosis take about a week for prophase I and about a month for the entire meiotic cycle. In females, prophase I begins long before a woman’s birth, during her early fetal development. Meiosis continues as much as decades later, during the monthly ovarian cycle, and is completed only after fertilization.
Independent Assortment
In addition to crossing over, meiosis provides a second source of genetic diversity. It is a matter of chance which member of a homologous pair goes to which daughter cell at anaphase I. For example, consider a diploid organism with two pairs of homologous chromosomes (pairs l and 2). One member of each pair came from the male parent of the organism (paternal 1 and 2), and the other came from the female parent (maternal 1 and 2). When cells in this organism undergo meiosis, a particular daughter nucleus could receive paternal 1 and maternal 2, paternal 2 and maternal 1, both maternal, or both paternal chromosomes. It all depends on how the homologous pairs line up at metaphase I. This phenomenon is called independent assortment.
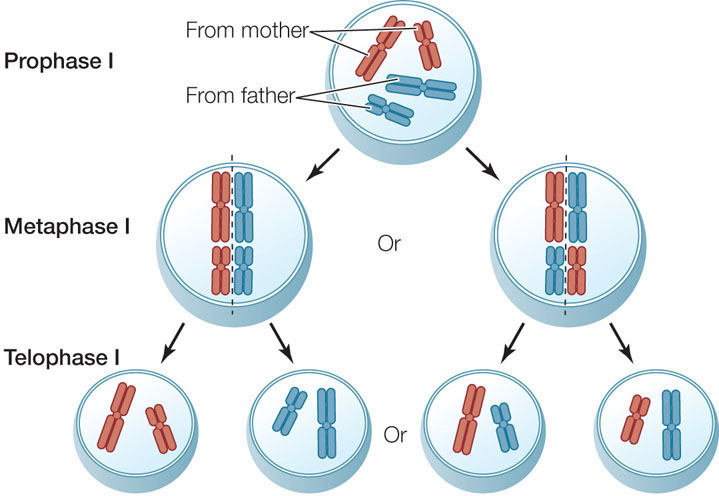
Note that of the four possible outcomes in the figure above, only two daughter nuclei receive either all maternal or all paternal chromosomes (apart from material exchanged by crossing over). The greater the number of chromosomes, the lower the probability of reestablishing the original parental combinations, and thus the greater the potential for genetic diversity. Most species of diploid organisms have more than two pairs of chromosomes. In humans, with 23 chromosome pairs, 223 (8,388,608) different combinations of maternal and paternal chromosomes can be produced just by the mechanism of independent assortment! Taking into account the extra genetic shuffling afforded by crossing over, the number of possible combinations is virtually infinite. Crossing over and independent assortment, along with the processes that result in mutations, provide the genetic diversity needed for evolution by natural selection.
145
We have seen how meiosis I is fundamentally different from mitosis. However, meiosis II is similar to mitosis in that it involves the separation of chromatids into daughter nuclei (see steps 7–11 in Figure 7.12). The final products of meiosis I and meiosis II are four haploid daughter cells, each with one set (n) of chromosomes.
Meiotic errors lead to abnormal chromosome structures and numbers
Meiosis is a complex process, and things occasionally go wrong. For example, chromosomes may break, homologs may fail to separate at anaphase I, or chromatids may fail to separate at anaphase II. The gametes formed from meiotic errors carry abnormal chromosomes, and when abnormal chromosomes take part in fertilization, the consequences for offspring can be significant.
Nondisjunction
Occasionally a homologous chromosome pair fails to separate (fails to “disjoin”) at anaphase I, or a pair of chromatids fail to separate at anaphase II. This failure to separate is referred to as nondisjunction. If a chromosome pair fails to separate at anaphase I, two of the four daughter nuclei will each end up with both members of that homologous pair, and the other two will have neither member of the pair. If nondisjunction occurs at anaphase II, only two of the four daughter nuclei will be affected: one will have an extra chromosome and the other will have one less than the full complement of chromosomes.
Using humans as an example, if during anaphase I the two homologs of chromosome 10 fail to separate, half the gametes will have two copies of chromosome 10, with a total of 24 chromosomes instead of 23. If one of these gametes fuses with a normal gamete during fertilization, the zygote will have 47 (23 + 24) chromosomes, with three copies of chromosome 10. The condition of having an abnormal number of chromosomes is called aneuploidy; having one extra chromosome is called trisomy, and missing one chromosome is called monosomy (FIGURE 7.14).
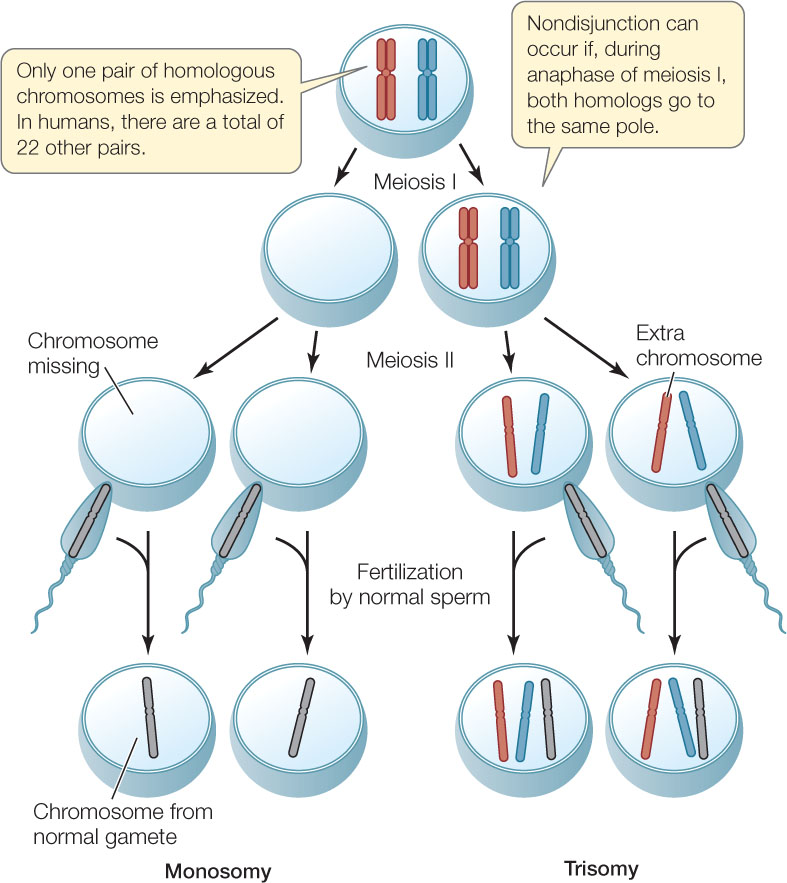
For reasons that are unclear, aneuploidy is a common and harmful condition in humans. About 10–30 percent of all conceptions show aneuploidy, but most of the embryos that develop from such zygotes do not survive to birth, and those that do often die before the age of 1 year. At least one-fifth of all recognized human pregnancies are spontaneously terminated (miscarried) during the first 2 months, largely because of trisomies and monosomies. The actual proportion of spontaneously terminated pregnancies is certainly higher, because the earliest ones often go unrecognized. The most common form of aneuploidy in humans is trisomy 16 (three copies of chromosome 16), but almost none of these embryos survive to birth. Among the few aneuploidies that allow survival is Down syndrome—trisomy 21. Such individuals generally have intellectual disabilities but can lead long and productive lives.
Polyploidy
Most organisms are either diploid (for example, most animals) or haploid (for example, most fungi). Under some circumstances, triploid (3n), tetraploid (4n), or higher-order polyploid nuclei may form. This can occur in a variety of ways. For example, there could be an extra round of DNA replication preceding meiosis, or there could be no spindle formed in meiosis II. Polyploidy occurs naturally in some animal species and in many plants.
LINK
Polyploidy can lead to reproductive isolation (the inability of two individuals to produce fertile offspring) and has probably led to speciation—the evolution of new species—as described in Concept 17.3
A diploid nucleus can undergo normal meiosis because there are two sets of chromosomes to make up homologous pairs, which separate during anaphase I. Similarly, a tetraploid nucleus has an even number of each kind of chromosome, so each chromosome can pair with its homolog. However, a triploid nucleus cannot undergo normal meiosis because one-third of the chromosomes would lack partners. Polyploidy has implications for agriculture, particularly in the production of hybrid plants. For example, ploidy (the number of chromosomes in the nucleus) must be taken into account in wheat breeding because there are diploid, tetraploid, and hexaploid wheat varieties. Polyploidy can be a desirable trait in crops and ornamental plants because it often leads to more robust plants with larger flowers, fruits, and seeds. In addition, triploid fruit varieties are desirable because they are infertile and therefore seedless.
146
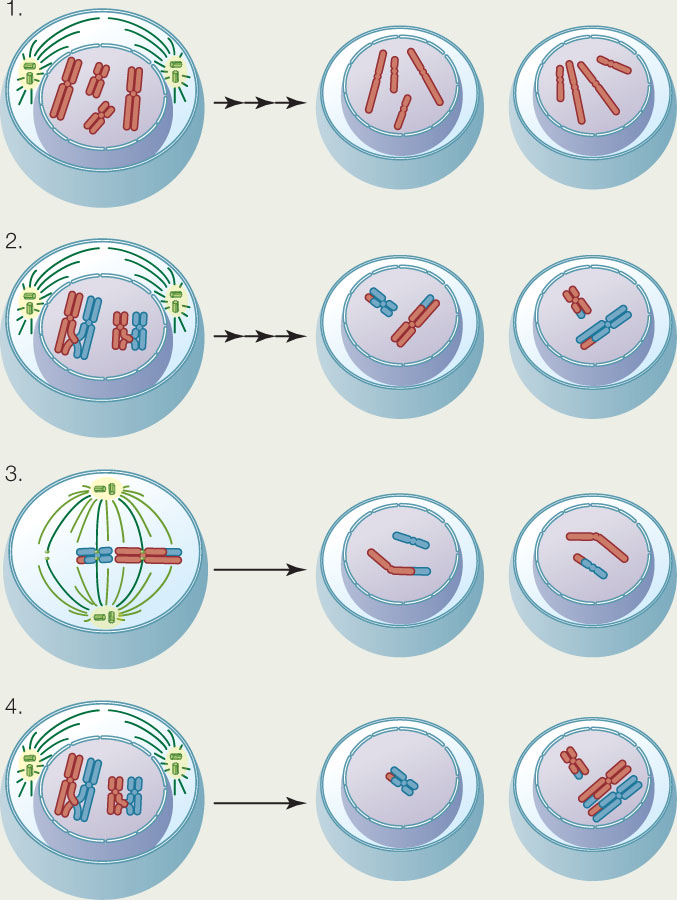
APPLY THE CONCEPT: Meiosis halves the nuclear chromosome content and generates diversity
Cells from a diploid organism (2n = 4) are shown undergoing division in the diagrams. For each diagram, indicate the type of cell division (mitosis or meiosis), the phase of division, and any special condition that is depicted.
Translocation
During crossing over in meiosis I, chromatids from homologous chromosome pairs break and rejoin. Occasionally this can happen between non-homologous chromosomes. The result is a translocation, and these are quite common, even in mitotic cells. As we will point out in our discussion of gene expression and its regulation in Chapters 10 and 11, the location of genes relative to other DNA sequences is important, and translocations can have profound effects on gene expression.
An example of a translocation known to occur in humans is a swap of material between chromosomes 9 and 22:
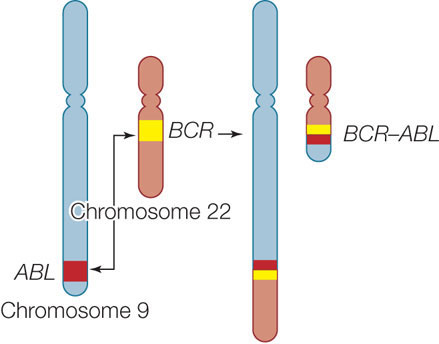
In this case, part of the BCR gene sequence on chromosome 22 comes to lie adjacent to part of the ABL gene sequence, which was translocated from chromosome 9. If the translocation occurs in a mitotic cell forming white blood cells, the result of this combination is a form of leukemia, a cancer of white blood cells.
A translocation that occurs during meiosis may be carried on the gametes that result and passed on to offspring at fertilization.
CHECKpoint CONCEPT 7.4
- How do crossing over and independent assortment during meiosis result in daughter nuclei that differ genetically?
- What are the differences between meiosis and mitosis?
- A vertebrate animal has a diploid number of 6. How many chromosomes are present in the following cells: A gamete? A gamete with monosomy of chromosome 2? A skin cell? A sperm cell at meiotic anaphase II?
An essential role of cell division in complex eukaryotes is to replace cells that die. What causes cells to die?