Concept 24.2: Apical Meristems Build the Primary Plant Body
As noted above, plants and animals differ in how they develop and function. Whereas animals use their mobility to forage for food, plants stay in place and grow toward resources, both above and below the ground. So plant shoots grow toward sunlight, and roots grow toward water and minerals in the soil. In most animals, growth is determinate: it ceases when the adult state is reached. Some plant organs, such as most leaves and flowers, show this characteristic. By contrast, the growth of roots and shoots is indeterminate, an open-ended process which can be lifelong.
527
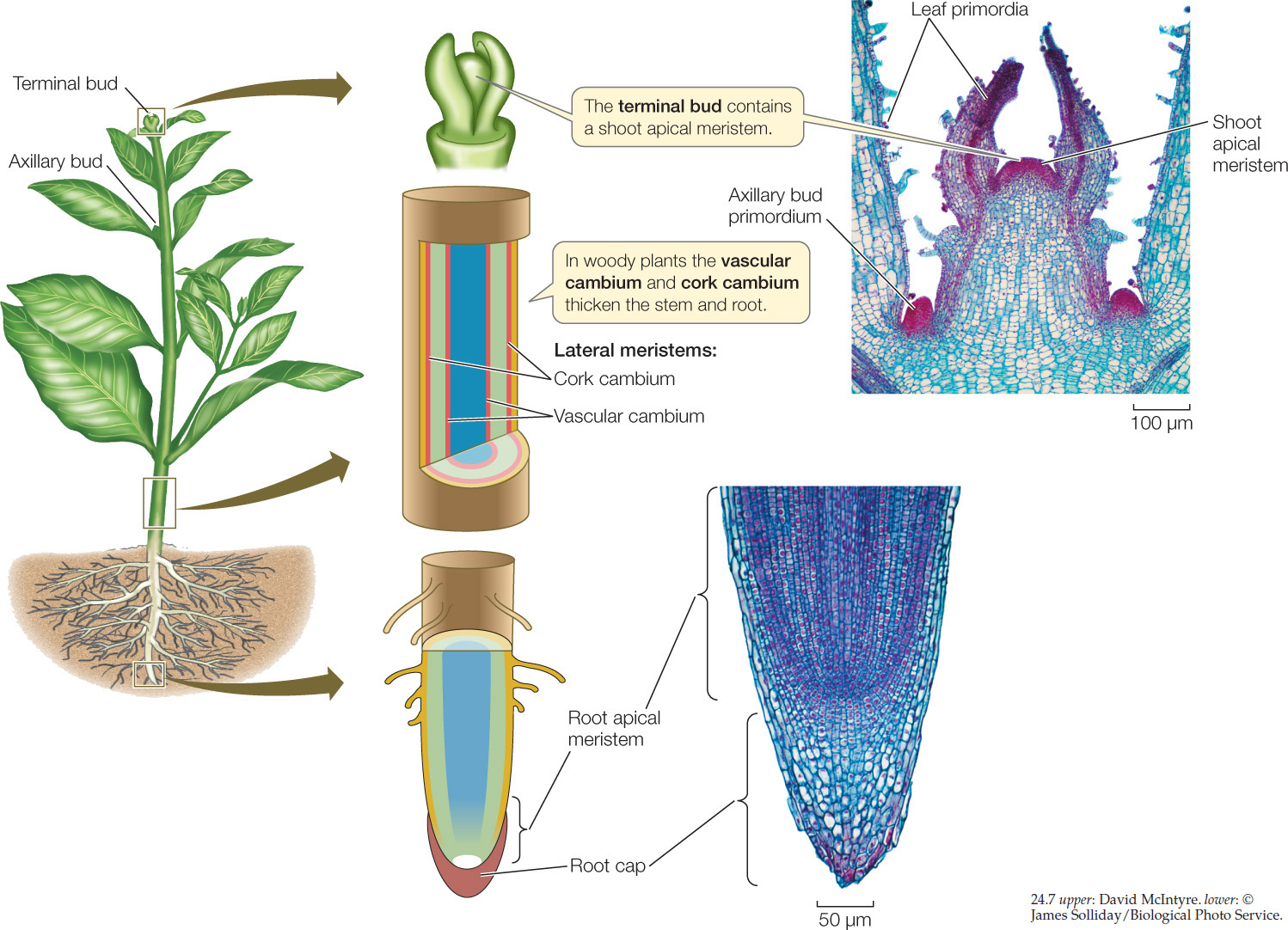
Plants grow in two ways (FIGURE 24.7):
- Primary growth results in the formation and lengthening of shoots and roots. Apical meristems produce primary growth, giving rise to the primary plant body. All seed plants have a primary plant body.
- Secondary growth increases plant thickness. Lateral meristems produce secondary growth, resulting in the secondary plant body. Secondary growth occurs in woody eudicots, such as trees and shrubs. Monocots typically do not undergo secondary growth.

Go to MEDIA CLIP 24.1 Rapid Growth of Brambles
PoL2e.com/mc24.1
The remainder of this section will focus on primary growth. We will return to secondary growth in Concept 24.3.
A hierarchy of meristems generates the plant body
Even before a seed germinates, the plant embryo inside the seed has two meristems: a shoot apical meristem at the end of the embryonic shoot and a root apical meristem near the end of the embryonic root (see Figure 24.5). The dividing cells of the meristems are comparable to animal stem cells: after their division, one daughter cell is capable of differentiating, and the other retains its undifferentiated phenotype.
The apical meristems form the root and shoot organs of the adult plant. They do so by producing three primary meristems.
From the outside to the inside of the root or shoot, which are both cylindrical, the primary meristems are the protoderm, the ground meristem, and the procambium. These meristems, in turn, give rise to the three tissue systems:
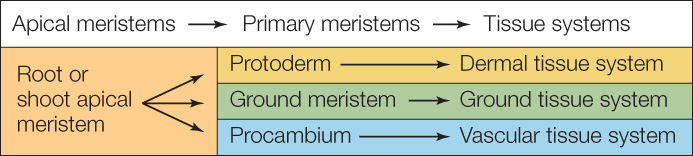
In plants that have secondary growth, the lateral meristems represent an additional level in the meristem hierarchy; for this reason, lateral meristems are also referred to as secondary meristems.
Not all meristems fit within this framework. The totipotency of many plant cells means that even differentiated, nonmeristematic cells can give rise to new meristems under certain circumstances. Examples include some forms of asexual reproduction and the propagation of plants from stem cuttings. The ability of nonmeristematic cells to further divide is important in wound repair.
528
The root apical meristem gives rise to the root cap and the root primary meristems
The root apical meristem produces all the cells that contribute to growth in the length of a root (FIGURE 24.8A). Some of the daughter cells from the apical (tip) end of the root apical meristem contribute to a root cap, which protects the delicate growing region of the root as it pushes through the soil. The root cap secretes a mucopolysaccharide (slime) that acts as a lubricant. Even so, the cells of the root cap are often damaged or scraped away and must therefore be replaced constantly. The root cap is also the structure that detects the pull of gravity and thus controls the downward growth of roots.
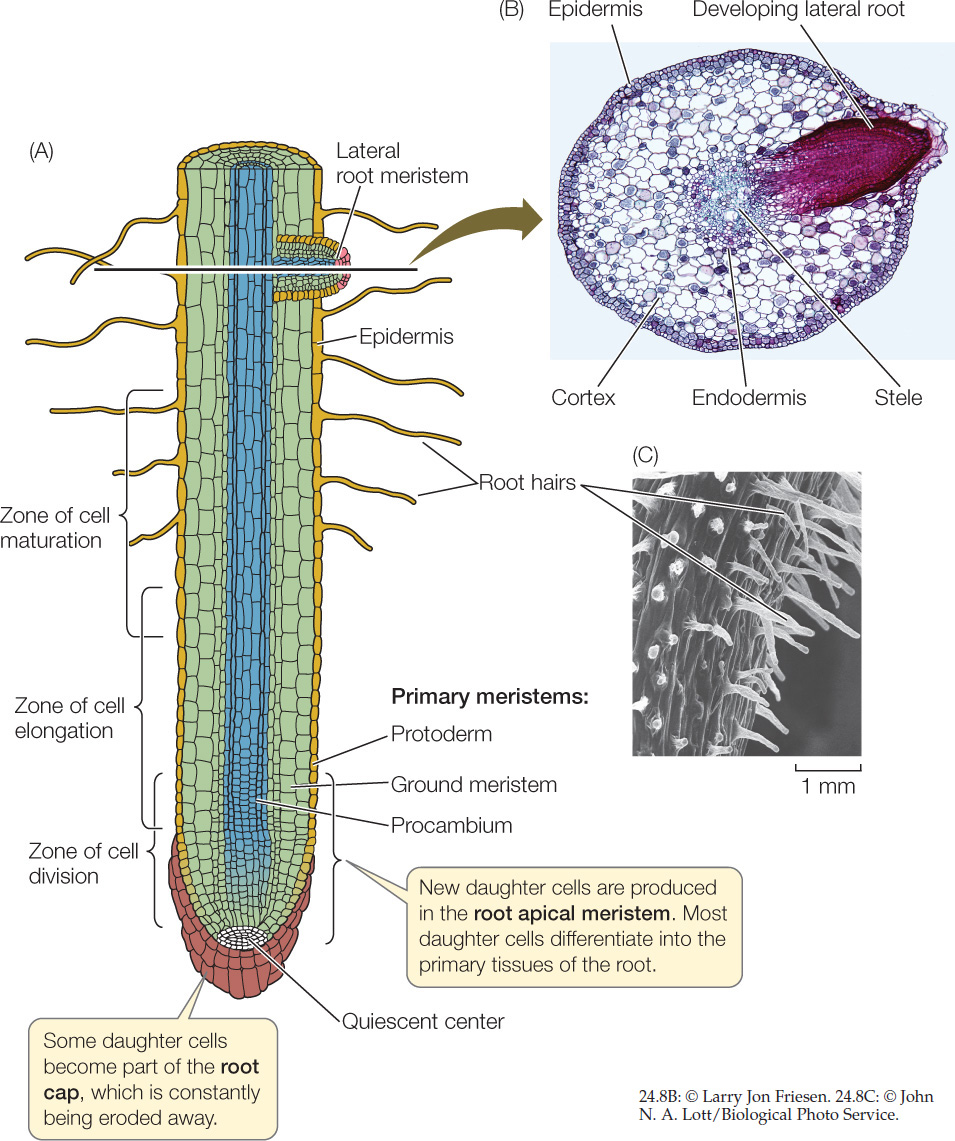
Above the root cap, the cells in the root tip are arranged in several zones. The apical and primary meristems constitute the zone of cell division, the source of all the cells of the root’s primary tissues. Daughter cells of the divisions of the apical meristem become the three cylindrical primary meristems: the protoderm, the ground meristem, and the procambium. These meristems in turn produce all the cells of the three tissues systems in the root. Just above this zone is the zone of cell elongation, where the newly formed cells are elongating and thus pushing the root farther into the soil. Above that zone is the zone of cell maturation, where the cells are completing their differentiation. These three zones grade imperceptibly into one another; there is no abrupt line of demarcation.
In the middle of the root apical meristem is a quiescent center, where cell divisions are rare. The rare divisions produce cells that ultimately become the root apical meristem.
The products of the root’s primary meristems become root tissues
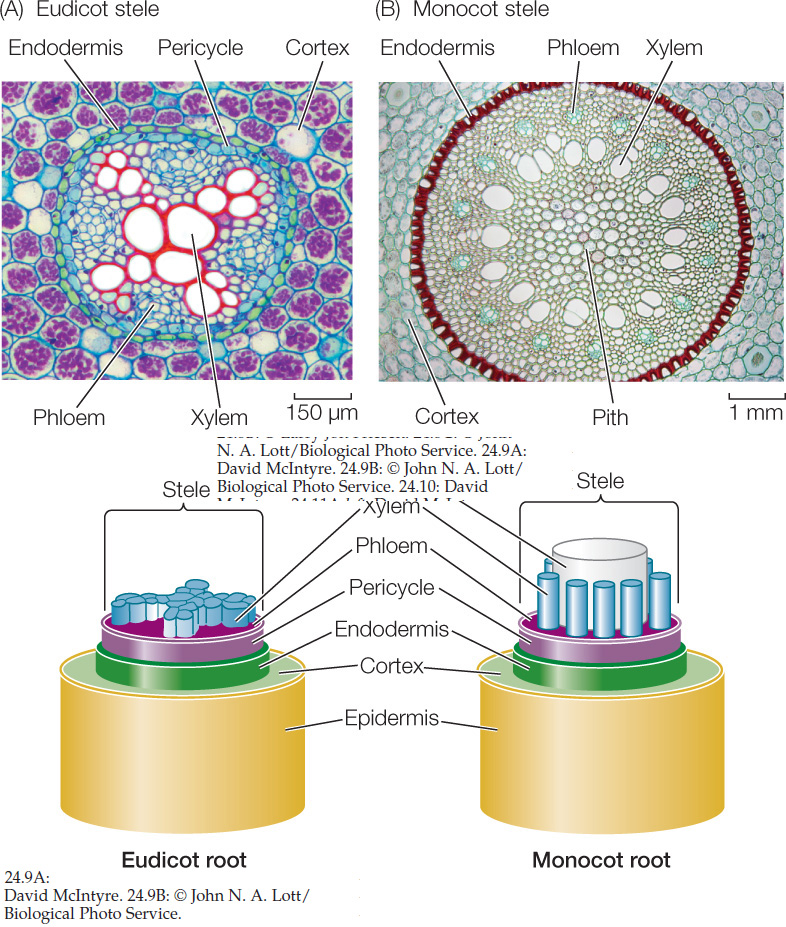
The products of the three primary meristems (the protoderm, ground meristem, and procambium) are the tissue systems of the mature root: the epidermis (dermal tissue), cortex (ground tissue), and stele (vascular tissue) (FIGURE 24.8B).
The protoderm gives rise to the epidermis, an outer layer of cells that protect the root and absorb mineral ions and water. Many of the epidermal cells produce long, delicate root hairs (FIGURE 24.8C).
Internal to the protoderm, the ground meristem gives rise to a region of ground tissue that is many cells thick, called the cortex. The parenchyma cells of the cortex are relatively unspecialized and often serve as storage depots. The innermost layer of the cortex is the endodermis (see Figure 24.8B). Unlike the cell walls of other cortical cells, those of the endodermal cells contain suberin, a waterproof substance. Strategic placement of suberin in only certain parts of the cell wall enables the cylindrical ring of endodermal cells to control the movement of water and dissolved mineral ions into the vascular tissue system.
LINK
The role of the endodermis is described in more detail in Concept 25.3
Interior to the endodermis is the vascular cylinder, or stele, produced by the procambium. The stele consists of three types of cells: pericycle, xylem, and phloem. The pericycle consists of one or more layers of parenchyma cells. It has three important functions:
- It is the tissue from which lateral roots arise.
- It can contribute to secondary growth by giving rise to lateral meristems that thicken the root.
- Its cells contain membrane transport proteins that export nutrient ions into cells of the xylem.
529
APPLY THE CONCEPT: Meristems build roots
A research team investigated how long root cells continued to divide after being produced by the root apical meristem. They incubated germinating bean seeds for 2 hours with their roots suspended in water containing radioactive thymidine, which is taken up by dividing cells and incorporated into their DNA as it replicates (making the DNA radioactive).
After this incubation period, the researchers removed some of the seeds (time 0 in the table) and took longitudinal slices from the roots (see Figure 24.8A). They transferred the rest of the seeds to water containing nonradioactive thymidine and left them there for various times (12, 36, and 72 hours) before taking root slices. They tallied the number of radioactive cells found in each of the four regions of each root and compared those numbers with the total number of cells in each region. The table shows the results.
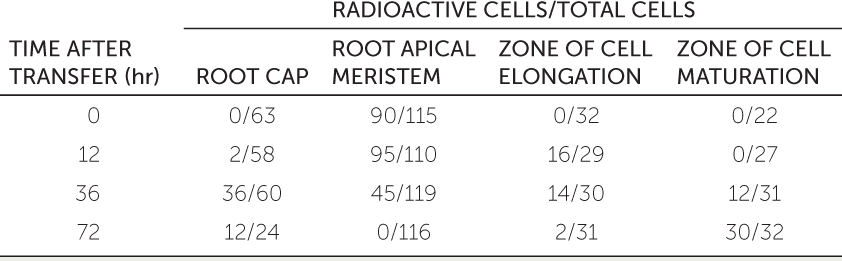
- At 0 hours, why did only root apical meristem cells contain radioactive thymidine?
- For each region, calculate the percentage of radioactive cells and plot that percentage over time.
- Explain the rise and fall in the percentage of radioactive cells in each region.
At the very center of the root of a eudicot lies the xylem. Seen in cross section, it typically has the shape of a star with a variable number of points (FIGURE 24.9A). Between the points are bundles of phloem. In monocots, a region of parenchyma cells called the pith typically lies in the center of the root, surrounded by xylem and phloem (FIGURE 24.9B). Pith, which often stores carbohydrate reserves, is also found in the stems of eudicots.
Go to ACTIVITY 24.1 Eudicot Root
PoL2e.com/ac24.1
Go to ACTIVITY 24.2 Monocot Root
PoL2e.com/ac24.2
The root system anchors the plant and takes up water and dissolved minerals
Water and mineral nutrients enter most plants through the root system, which is located in the soil. Although hidden from view, the root system is often larger than the visible shoot system. For example, the root system of a 4-month-old winter rye plant (Secale cereale) was found to be 130 times larger than the shoot system, with almost 13 million branches that had a cumulative length of over 500 kilometers! The large size of root systems gives them an enormous surface area for absorbing water and nutrients from the soil.
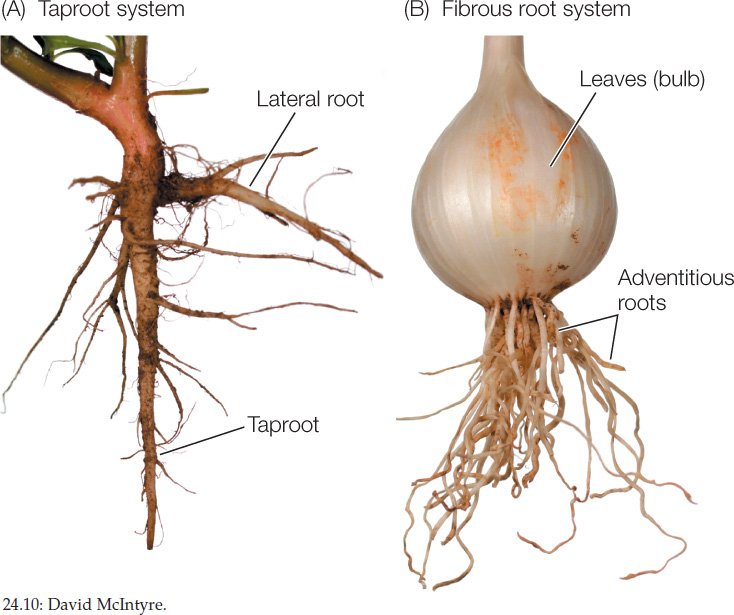
The root system of angiosperms originates in an embryonic root called the radicle. In most eudicots the radicle develops as a primary root (called the taproot) that extends downward by tip growth and outward by initiating lateral roots (FIGURE 24.10A).
In monocots, the radicle-derived initial root is short-lived. Monocots form a fibrous root system composed of numerous thin roots that are all roughly equal in diameter (FIGURE 24.10B). These roots are adventitious: they arise from stem tissues above the initial root. Fibrous root systems often form dense networks of roots near the soil surface, binding the soil together. The fibrous root systems of grasses, for example, may protect steep hillsides where runoff from rain would otherwise cause erosion.
530
The products of the shoot’s primary meristems become shoot tissues
The shoot apical meristem, like the root apical meristem, forms three primary meristems: protoderm, ground meristem, and procambium. These primary meristems, in turn, give rise to the three shoot tissue systems. The shoot apical meristem repetitively lays down the beginnings of leaves and axillary buds (corresponding to the phytomers in Figure 24.1). Leaves arise from bulges called leaf primordia, which form as cells divide on the sides of the shoot apical meristem (see Figure 24.7). Bud primordia form above the bases of the leaf primordia, where they may initiate new shoots. The growing stem has no protective structure analogous to the root cap, but the leaf primordia can act as a protective covering for the shoot apical meristem.
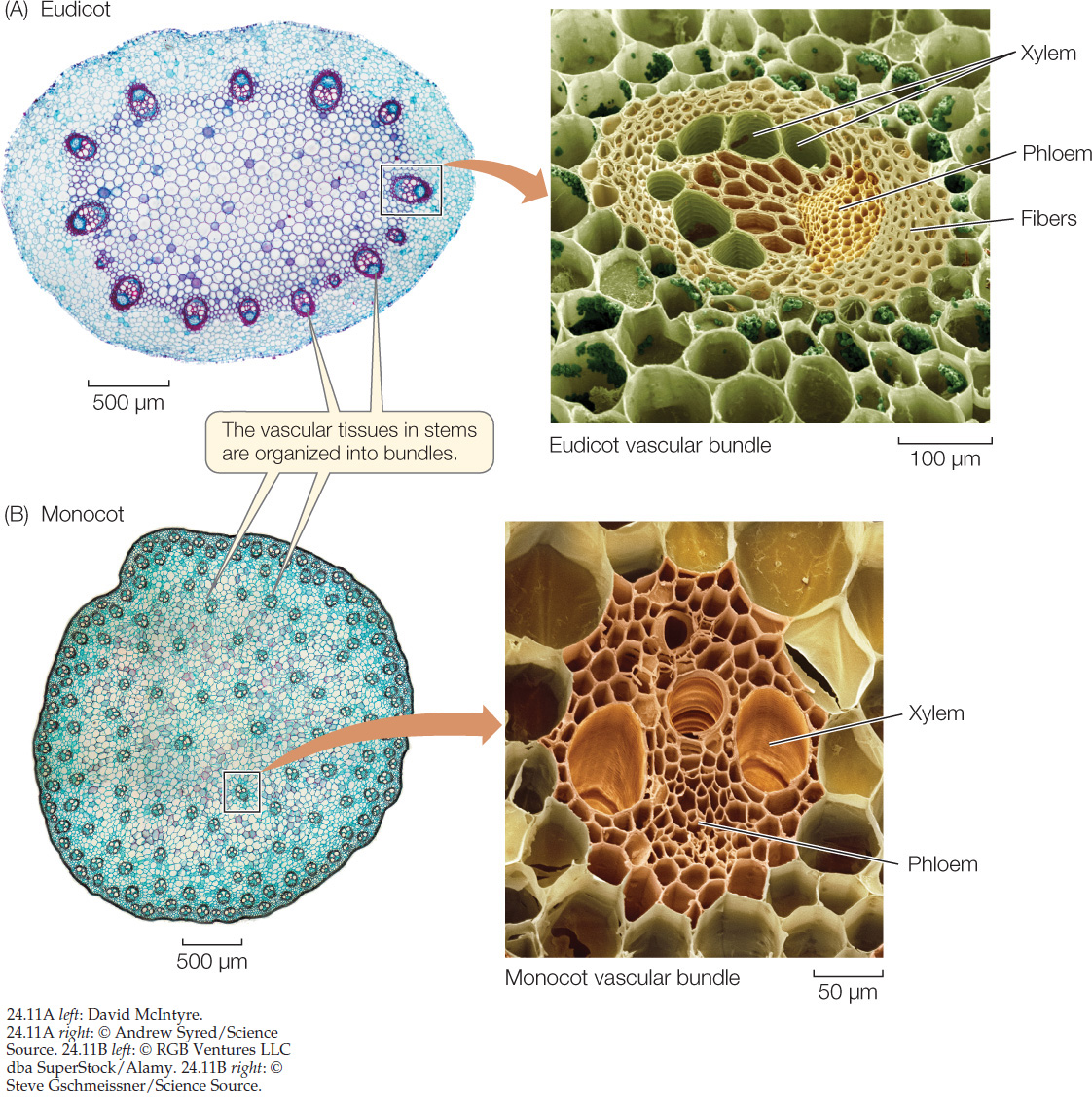
As in roots, the shoot protoderm, ground meristem, and procambium give rise, respectively, to the shoot epidermis, the shoot cortex, and the shoot vascular system. The vascular tissue of young stems is divided into discrete vascular bundles (FIGURE 24.11). Each vascular bundle contains both xylem and phloem. In eudicots, the vascular bundles are arranged along the periphery and appear in a circle in cross section, but in monocots they are scattered throughout the stem.
Go to ACTIVITY 24.3 Eudicot Stem
PoL2e.com/ac24.3
Go to ACTIVITY 24.4 Monocot Stem
PoL2e.com/ac24.4
531
Leaves are photosynthetic organs produced by shoot apical meristems
For most of its life, a plant produces leaves from shoot apical meristems. As shown in Figure 24.7, leaves originate from leaf primordia at the sides of the shoot apical meristem. Unlike the growth of roots and stems, the growth of leaves is finite, or determinate: leaves generally stop growing once they reach a predetermined mature size and shape.
Typically, the blade of a leaf is a thin, flat structure. In many eudicots, the blade is attached to the stem by a stalk called a petiole (see Figure 24.1). Often the petiole can adjust the orientation of the leaf blade relative to the rays of the sun, ranging from perpendicular to the light rays (to maximize photosynthesis) to parallel (to prevent overheating). Some leaves track the sun over the course of the day, moving so that they constantly face it. Monocots typically do not have petioles, and the base of the leaf may form a sheath that wraps around the stem.
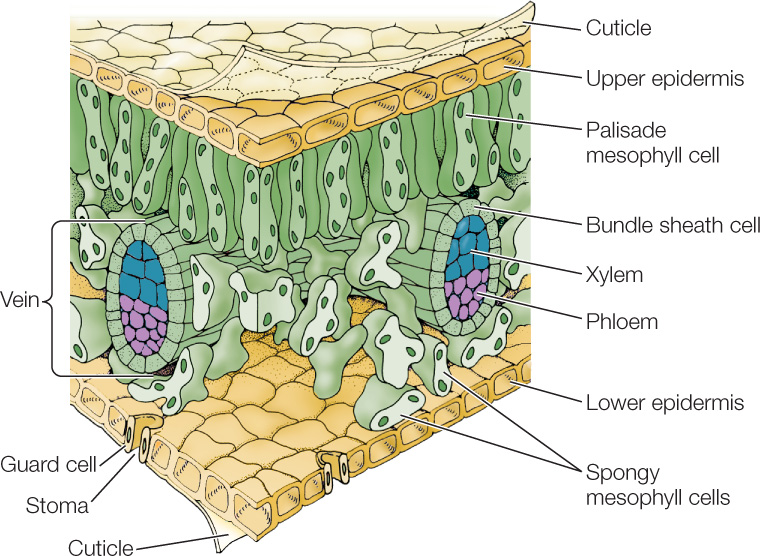
Leaf anatomy is beautifully adapted to carry out photosynthesis and to support that process by exchanging the gases O2 and CO2 with the environment, limiting evaporative water loss, and exporting the products of photosynthesis to the rest of the plant. FIGURE 24.12 shows a section of a typical eudicot leaf in three dimensions.
Go to ACTIVITY 24.5 Eudicot Leaf
PoL2e.com/ac24.5
Most eudicot leaves have two zones of photosynthetic parenchyma tissue called mesophyll (which means “middle of the leaf”). The upper layer or layers of mesophyll, which consist of elongated cells, constitute a zone called palisade mesophyll. The lower layer or layers, which consist of irregularly shaped cells, constitute a zone called spongy mesophyll. Within the mesophyll is a network of air spaces through which CO2 can diffuse to photosynthesizing cells. Vascular tissue branches extensively throughout the leaf, forming a network of veins that extend to within a few cell diameters of all its cells, ensuring that the mesophyll cells are well supplied with water and mineral nutrients by the xylem. The products of photosynthesis are loaded into the phloem of the veins for export to the rest of the plant.
LINK
Photosynthesis is described in Concepts 6.5 and 6.6
Covering virtually the entire leaf on both its upper and lower surfaces is a layer of nonphotosynthetic epidermal cells. These cells secrete a cuticle that is impermeable to water. Although the epidermis keeps water in the leaf, it also keeps out CO2—which is needed for photosynthesis. Therefore water and gases are exchanged through pores in the leaf called stomata (see Concept 25.3).
Plant organs can have alternative forms and functions
Flowering plants are incredibly diverse in form. Leaves, stems, and roots of many shapes can perform the basic functions of plant life: photosynthesis, the acquisition of water and nutrients, sensing the environment, and reproduction. These plant organs may also be modified to serve more specialized functions.
Taproots, for example, often function as nutrient storage organs, as in carrots, sugar beets, and sweet potato (FIGURE 24.13A). In some monocots—corn and some palms, for example—adventitious roots function as props to help support the shoot (FIGURE 24.13B). Prop roots are critical to these plants, which, unlike most eudicot tree species, are unable to support aboveground growth through the thickening of their stems.
Various modifications of stems are also seen in nature. The tuber of a potato, for example—the part of the plant eaten by humans—is not a root but rather an underground stem. The “eyes” of a potato are depressions containing axillary buds—in other words, a sprouting potato is just a branching stem (FIGURE 24.13C). Many desert plants have enlarged, water-retaining stems (FIGURE 24.13D). The runners of strawberry plants are horizontal stems from which roots grow at frequent intervals (FIGURE 24.13E). If the links between the rooted portions are broken, independent plants can develop on each side of the break—a form of vegetative (asexual) reproduction (see Concept 7.1).
Leaves are sometimes highly modified for special functions. For example, some leaves serve as storage depots for energy-rich molecules, as in the bulbs of onions. In succulents, the leaves store water. The protective spines of cacti are modified leaves (see Figure 24.13D). Other plants have modified portions of leaves called tendrils that support the plant by wrapping around other structures or plants (FIGURE 24.13F).

These are all examples of vegetative plant organs whose functions deviate from the norm. The evolution of these novel roles for established organs is an example of natural selection working with what is already present, as well as an example of the interaction between evolution and development—the topic of evolutionary developmental biology (evo-devo).
532
LINK
See Concepts 14.4 and 14.5 for a discussion of evolutionary developmental biology
In Concept 24.4 we will return to the modification of plant organs—in this case, the ways they have been manipulated by artificial selection to meet the needs of humans.
CHECKpoint CONCEPT 24.2
- What is the difference between determinate and indeterminate growth? What plant organs undergo each type of growth?
- What are the selective advantages of indeterminate growth?
Primary growth forms the plant organs and lengthens them. We now turn to secondary growth, in which some organs can grow in thickness.
533